Firefly bioluminescence is a remarkably efficient chemical reaction. Nearly 100% of the energy is released as light rather than heat. Here’s what’s happening at the molecular level:
The core reaction
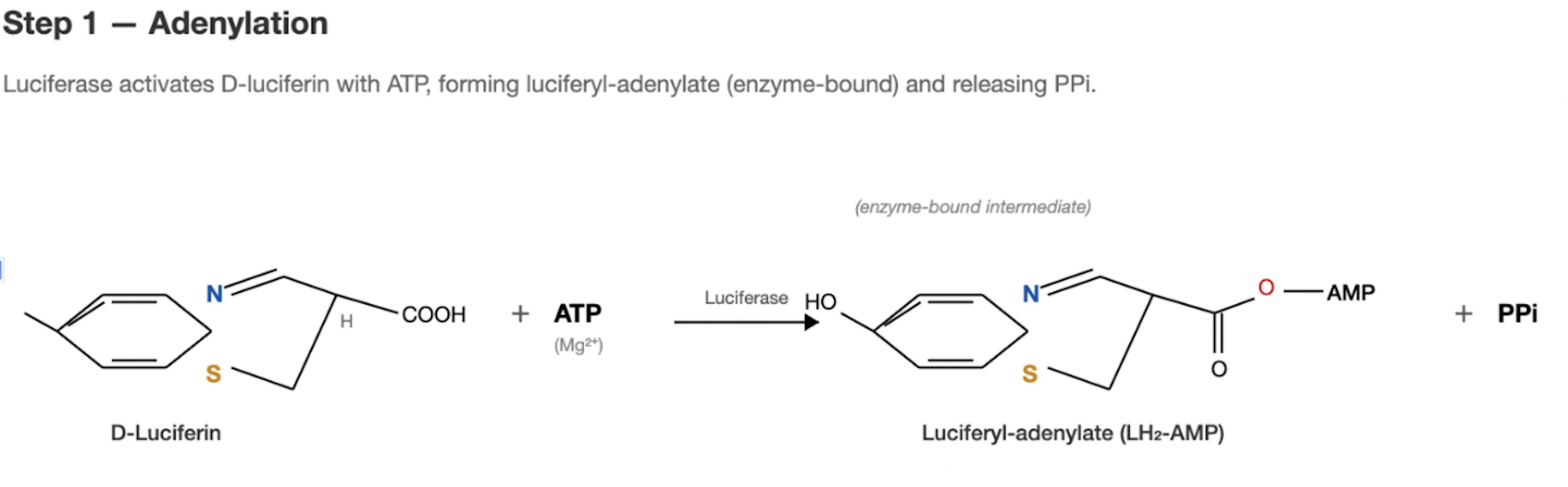
Step 1: Activation
Luciferase (the enzyme) catalyzes a two-step reaction (Conti et al. 1996; Inouye 2010). First it binds D-luciferin and ATP (with Mg²⁺ as a cofactor) forming luciferyl-adenylate (luciferin-AMP) and releasing pyrophosphate (PPi). This is essentially “charging up” the substrate.
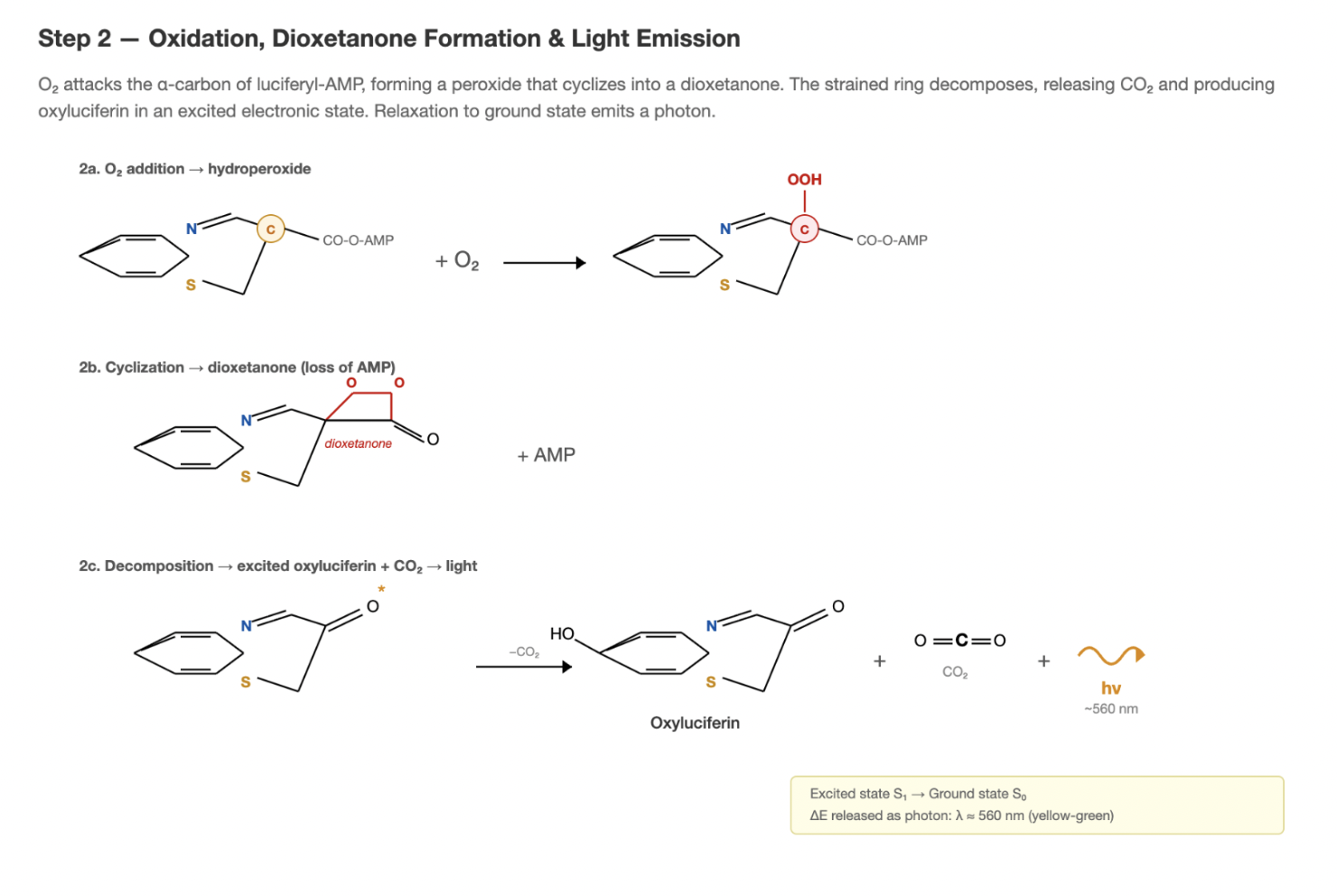
Step 2: Oxidation and light emission
The luciferyl-adenylate intermediate then reacts with molecular oxygen (O₂) (Conti et al. 1996). This oxidation produces a highly unstable dioxetanone ring intermediate, which spontaneously decomposes to yield oxyluciferin, CO₂, and AMP and a photon of light (~560 nm, yellow-green).
This light comes from oxyluciferin being produced in an electronically excited state and then relaxing to its ground state by emitting that photon.
Dictionary (6 terms)
- Luciferase
- The enzyme that catalyzes the light-producing reaction.
- D-luciferin
- The small molecule "fuel" that luciferase uses to generate light.
- Luciferyl-adenylate (LH₂-AMP)
- A short-lived activated form of luciferin created inside the enzyme before oxygen reacts.
- PPi (pyrophosphate)
- A common cellular byproduct released when ATP is used.
- Dioxetanone
- A very unstable high-energy intermediate whose breakdown ultimately leads to light emission.
- Oxyluciferin
- The oxidized "spent" form of luciferin left after the flash.
The luciferin biosynthesis problem
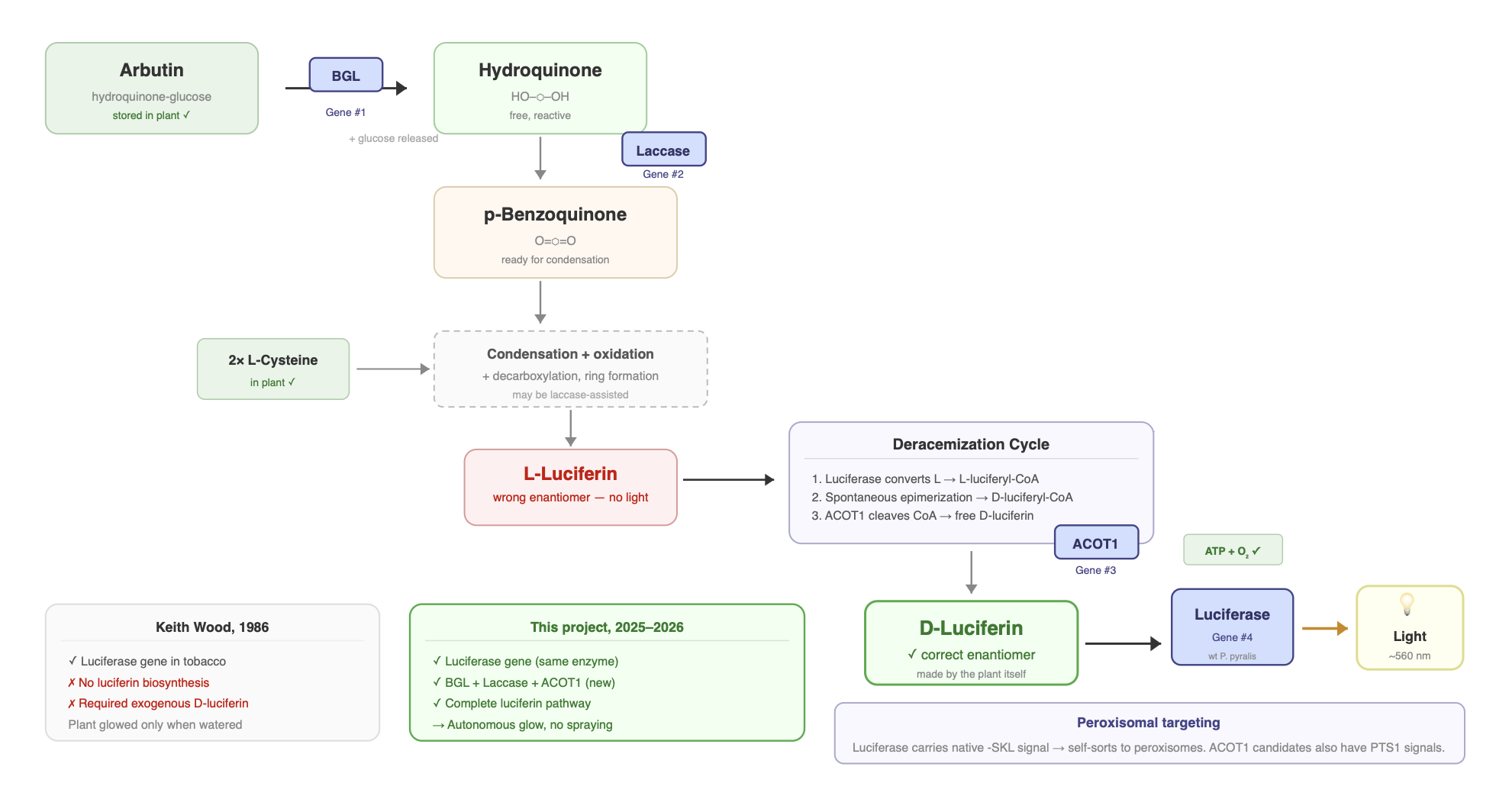
This is where it gets really interesting and where the field has been stuck for decades. Keith Wood showed in 1986 that if you put the luciferase gene into plants and spray them with exogenous luciferin, they glow (Ow et al. 1986). The luciferase cDNA had been cloned just a year earlier (de Wet et al. 1985). But that’s not autonomous, you need to continuously supply the substrate. The question is: where does D-luciferin come from in the firefly? The pathway has been progressively worked out over the last decade, but certain parts remain unclear.
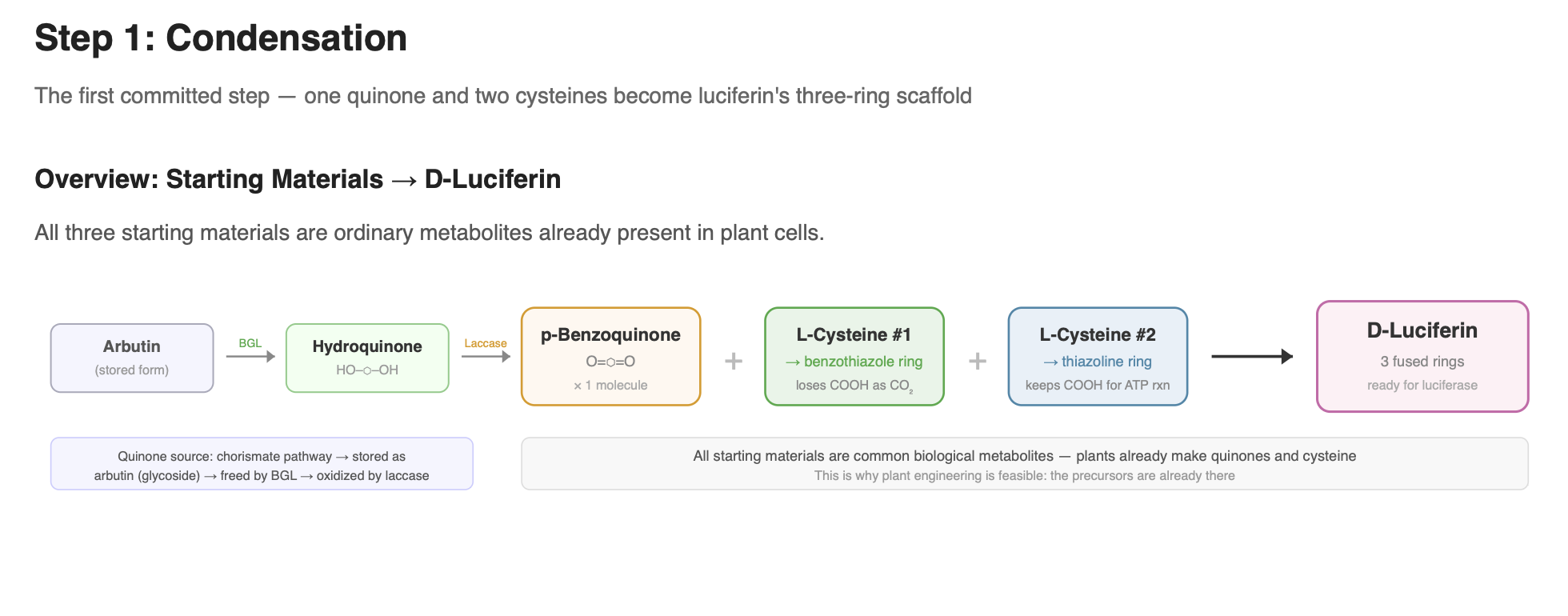
Step 1: The condensation of the quinone with L-cysteine
That’s the first moment where something luciferin-specific is happening. Everything before it is just general metabolism making normal molecules. The condensation is where the firefly’s unique chemistry begins, it’s the first committed step toward building luciferin.
Current evidence supports that luciferin forms from one molecule of a quinone and two molecules of L-cysteine (Oba et al. 2013). These are ordinary biological metabolites: quinone/hydroquinone redox systems are widespread in cells (CoQ10 being a familiar example), and cysteine is a standard amino acid present in all tissues. This makes the chemistry particularly relevant for engineering efforts, since the precursors already exist in plants.
Hydroquinone → quinone (activation step)
Before cysteine can attach, hydroquinone must be oxidized into its reactive quinone form. Quinones are much more chemically reactive than hydroquinones and readily participate in the condensation reactions that begin building the luciferin scaffold. This oxidation can occur spontaneously in air, but in living systems it is likely assisted by oxidase enzymes. This is the step that produces the electrophilic quinone that allows cysteine to attack and begin forming the benzothiazole core.
The first cysteine reacts with the quinone to form the benzothiazole core via a 2-S-cysteinylhydroquinone intermediate (Kanie et al. 2018), and during this step its carboxyl group is lost as CO₂ through decarboxylation. A second cysteine then condenses with this intermediate to form the thiazoline ring and contributes the carboxylate group required for the ATP-dependent luciferase reaction. Together, these steps generate luciferin’s characteristic heterocyclic structure.
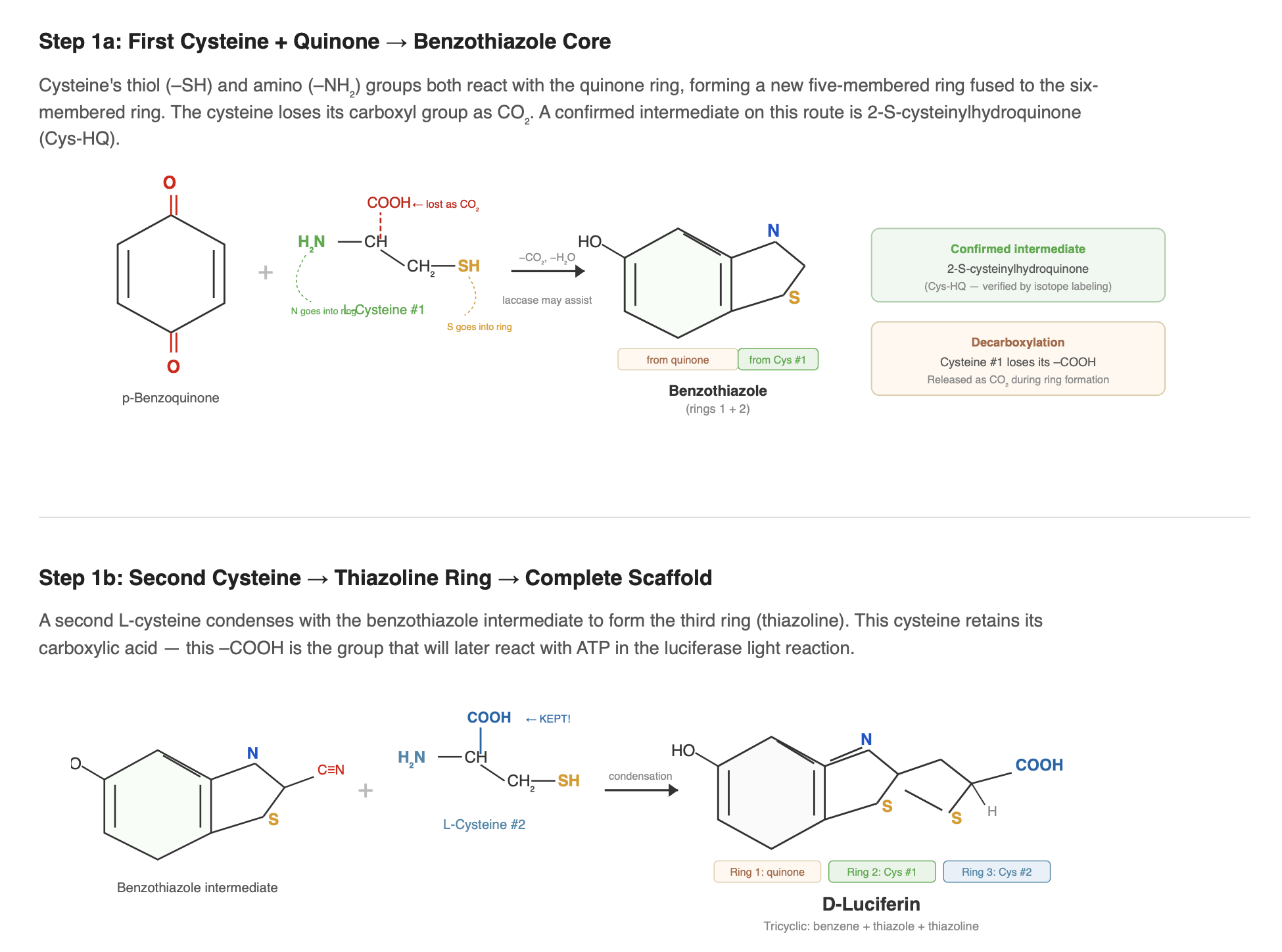
This condensation chemistry can occur non-enzymatically: p-benzoquinone mixed with cysteine in a neutral buffer produces luciferin at low yield (Kanie et al. 2016). In the firefly, however, the process is likely enzyme-assisted. Copper oxidases such as laccase enhance luciferin formation from hydroquinone and cysteine in vitro (de Souza et al. 2022), suggesting that regulated quinone redox chemistry may play a role in vivo. Hydroquinone itself is thought to be stored in a protected glycosylated form, possibly as arbutin, and released by β-glucosidase when needed (Oba et al. 2013).
The system functionally depends on D-luciferin, since it is the only enantiomer efficiently used by luciferase to produce light.

Dictionary (19 terms)
- Condensation
- A reaction where two smaller molecules join together to form one larger molecule. When they join, a tiny molecule such as water is usually released as a byproduct.
- Quinone
- A small ring-shaped molecule that cells commonly use to move electrons around. It exists because cells constantly need to transfer energy and electrons during metabolism. Quinones are good at accepting and giving away electrons, which makes them useful in energy systems like respiration and photosynthesis.
- Hydroquinone
- The reduced form of a quinone. In simple terms, it is the same ring structure but holding extra electrons. Cells can switch quinone and hydroquinone back and forth depending on whether they need to carry or release energy.
- Redox pair
- Two versions of the same molecule that can convert into each other by gaining or losing electrons. One form holds more energy, the other holds less. Quinone and hydroquinone are a classic example.
- L-cysteine
- One of the twenty normal amino acids used to build proteins. It is special because it contains sulfur, which makes it chemically reactive and useful for building more complex molecules.
- Ubiquinone (Coenzyme Q10)
- A quinone used by cells to move electrons during energy production inside mitochondria. It is present in almost all plants and animals. Yes, this is the same CoQ10 that is sold as a supplement.
- Benzothiazole
- A structure made from two connected rings. One is a standard benzene ring and the other is a smaller ring that contains sulfur and nitrogen. This type of structure is common in many natural molecules and drugs.
- Decarboxylation
- A reaction where a small piece of a molecule called a carboxyl group is removed and released as carbon dioxide. This often happens when molecules are being reshaped into new structures.
- Thiazoline
- A small five-membered ring that contains sulfur and nitrogen. It is not fully stabilized like an aromatic ring. In luciferin, this ring carries the acid group that lets the molecule react with ATP.
- Tricyclic
- A molecule made of three connected rings.
- Laccase
- An enzyme found in many plants and fungi that helps with oxidation reactions. It uses copper to pull electrons away from other molecules. In nature, laccases are often used to modify or break down complex plant chemicals like lignin.
- β-Glucosidase (BGL)
- An enzyme that cuts a sugar off of another molecule. Organisms often attach sugars to chemicals to store them safely. This enzyme removes the sugar and releases the active molecule when it is needed.
- Arbutin
- A storage form of hydroquinone where a glucose sugar is attached to it. This makes hydroquinone safer and less reactive. When the organism needs it, the sugar can be removed to release hydroquinone.
- Glycoside
- A molecule where a sugar is attached to another chemical. Cells often use this as a storage strategy. The sugar keeps the molecule stable and non-reactive until an enzyme removes it.
- In vitro
- Something studied in a controlled lab setting, such as in a test tube or dish, rather than inside a living organism.
- Stereochemistry
- The three-dimensional arrangement of atoms in a molecule. Some molecules come in two mirror-image forms that look the same on paper but behave differently in biology. These are called D and L forms.
- Enantioselective
- A reaction that prefers to make one mirror-image version of a molecule more than the other.
- Racemase
- An enzyme that can flip one mirror-image form of a molecule into the other.
- Kinetic resolution
- When an enzyme strongly prefers to use one mirror-image version of a molecule and ignores the other. Over time, this makes one form dominate.
Step 2: Oxidation: benzothiazoline to benzothiazole
At some point during the pathway, the benzothiazoline ring must be oxidized to become a benzothiazole, the five-membered ring becomes fully aromatic. In benzothiazoline, a C–H bond in the five-membered ring interrupts the continuous loop of conjugated double bonds. Oxidation removes two hydrogens and forms a new double bond, completing an unbroken ring of delocalized electrons. Think of it like a highway with a gap. Once the gap is filled, electron traffic flows smoothly around the entire ring. The molecule becomes significantly more stable.
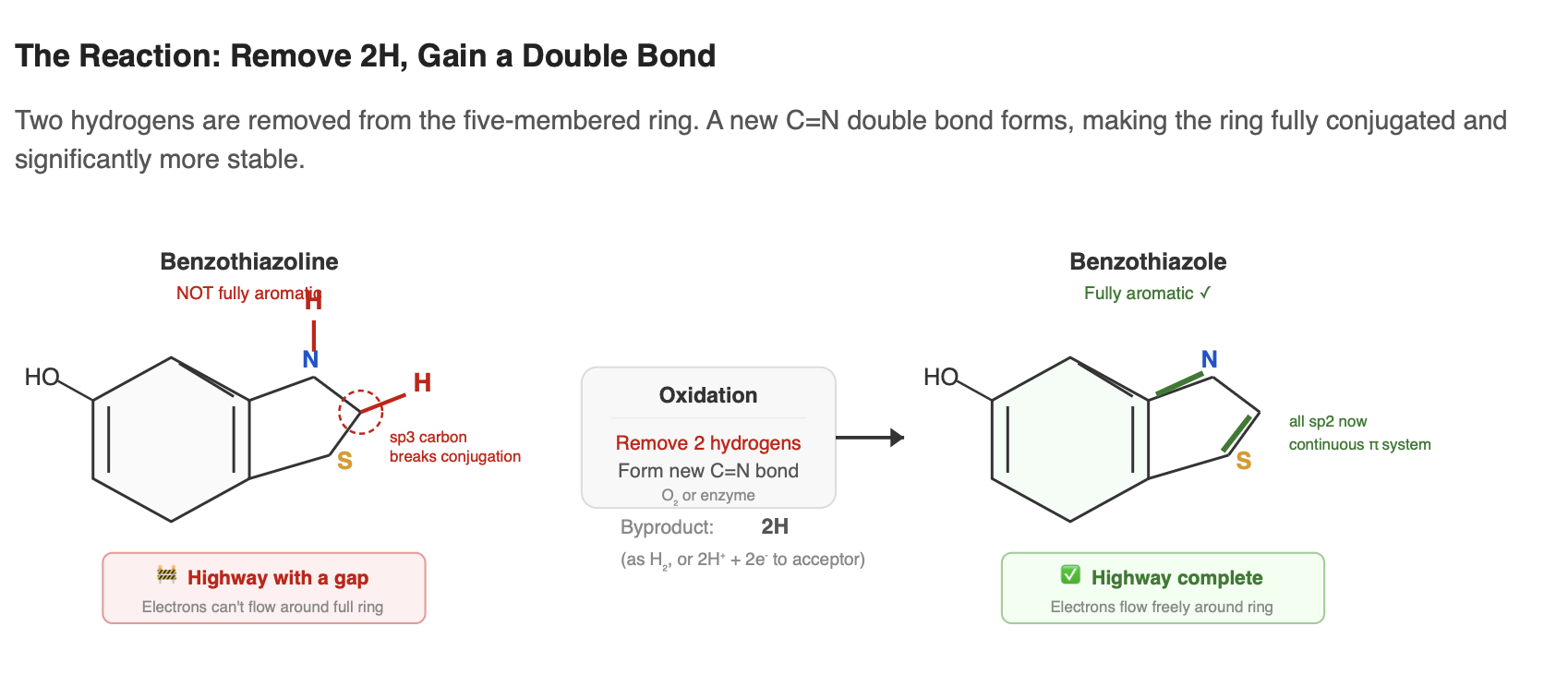
This oxidation can happen spontaneously. Benzothiazolines are prone to air oxidation, and in the non-enzymatic one-pot reaction (benzoquinone + cysteine in neutral buffer), molecular oxygen alone is sufficient (Kanie et al. 2016). In fireflies, however, the step is likely enzyme-assisted to control rate, timing, and selectivity. The responsible enzyme has not been definitively identified. Candidates include a dedicated oxidase, a laccase already involved in earlier steps of the pathway, or a side activity of another pathway enzyme.
Notably, the exact ordering of this oxidation relative to addition of the second cysteine remains unclear. The benzothiazoline could be oxidized before the second condensation step, or the fully assembled scaffold could be oxidized later as a finishing reaction. The in-vivo sequence has not been established.
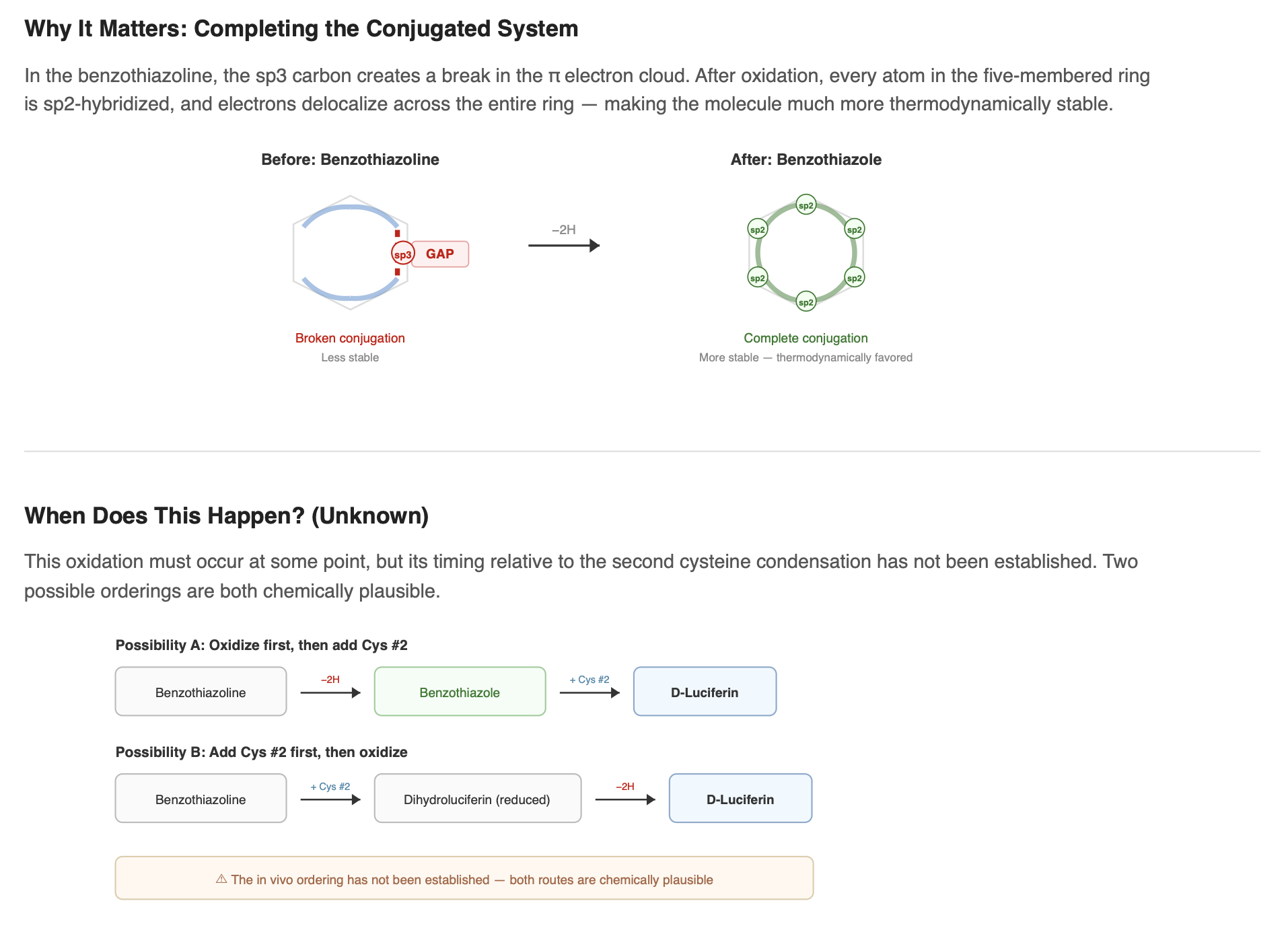
What else must happen
After the benzothiazole core is formed and the second cysteine is added, several additional modifications must occur to produce the final, functional D-luciferin. The exact ordering and enzymes involved remain uncertain, but here is what we know about each.
The 6-hydroxyl group
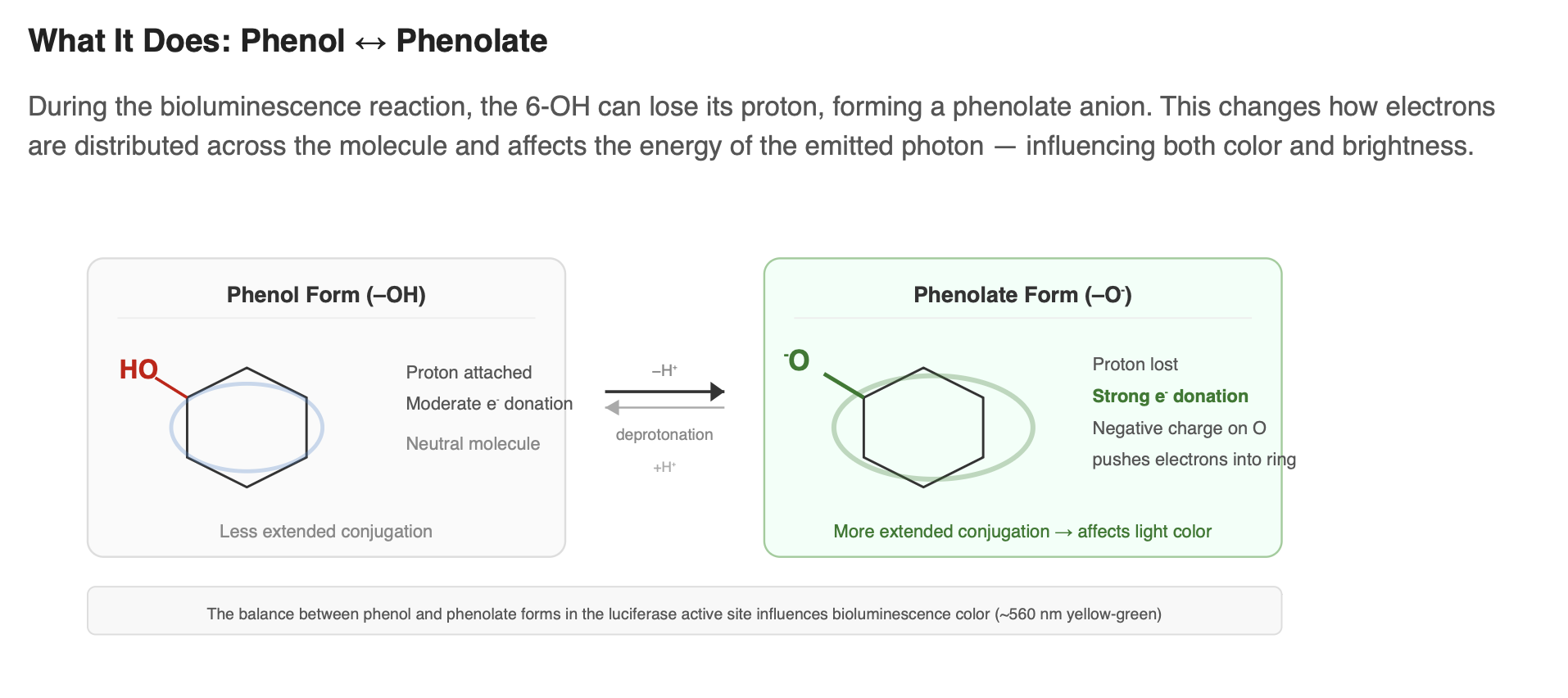
D-luciferin contains a hydroxyl group (–OH) at the 6-position of the benzothiazole ring. This group plays an important role in efficient bioluminescence. It participates in the molecule’s conjugated system and helps shape how electrons are distributed across the structure. During the light-producing reaction, the –OH can lose its proton to form a phenolate (–O⁻). This change affects the energy of the excited state and is known to influence both the color and efficiency of light emission.
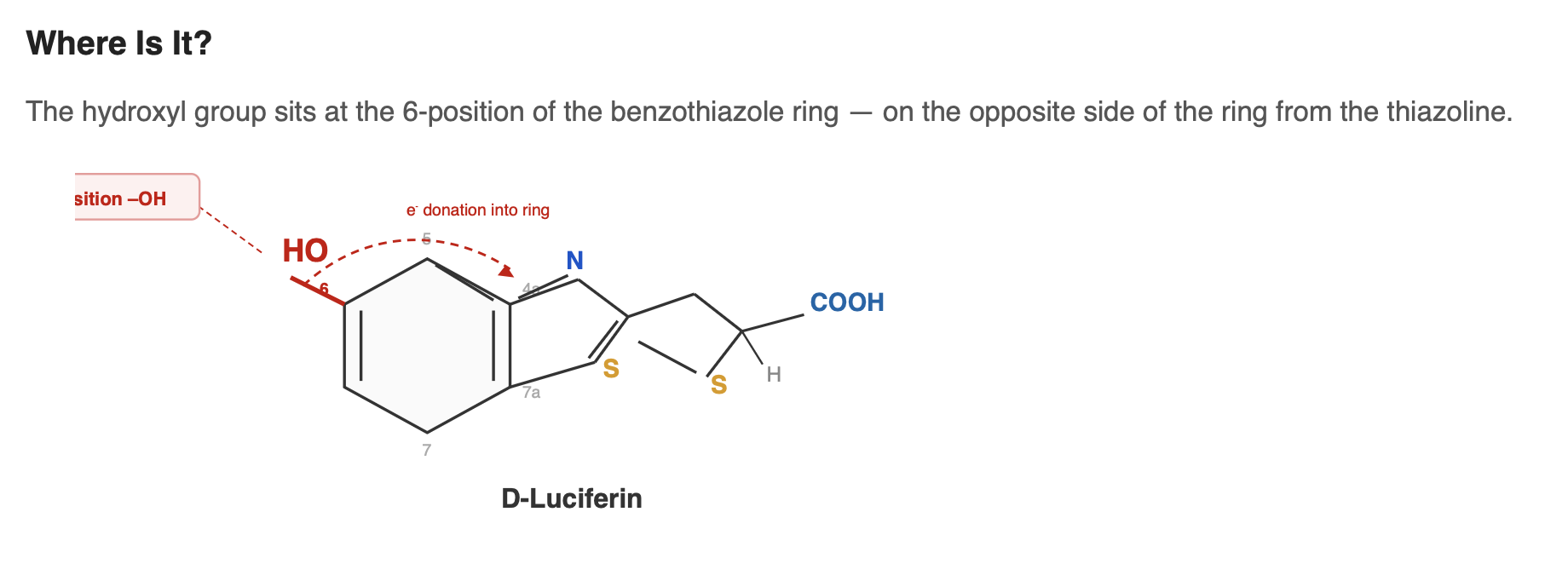
Where this hydroxyl group comes from in biosynthesis is still an open question. If the pathway begins with a precursor like p-benzoquinone, the oxygen atom may already be positioned correctly and carried through into the final structure, meaning no separate hydroxylation step would be needed. If the precursor does not provide the correct hydroxyl pattern, then an additional enzyme would likely be required to install it.
Stereochemical inversion: L-luciferin to D-luciferin
This is one of the most interesting unsolved problems in the pathway. Both cysteine molecules entering biosynthesis are L-cysteine, the natural form found in all living organisms. Yet efficient light production depends on D-luciferin, because firefly luciferase uses the D-enantiomer far more effectively than the L form.

The current leading model, first proposed by Niwa et al. (2006) and later validated experimentally by Zhang et al. (2020), proposes that the biosynthetic pathway initially produces L-luciferin, which is then converted to D-luciferin through a three-step deracemization process:
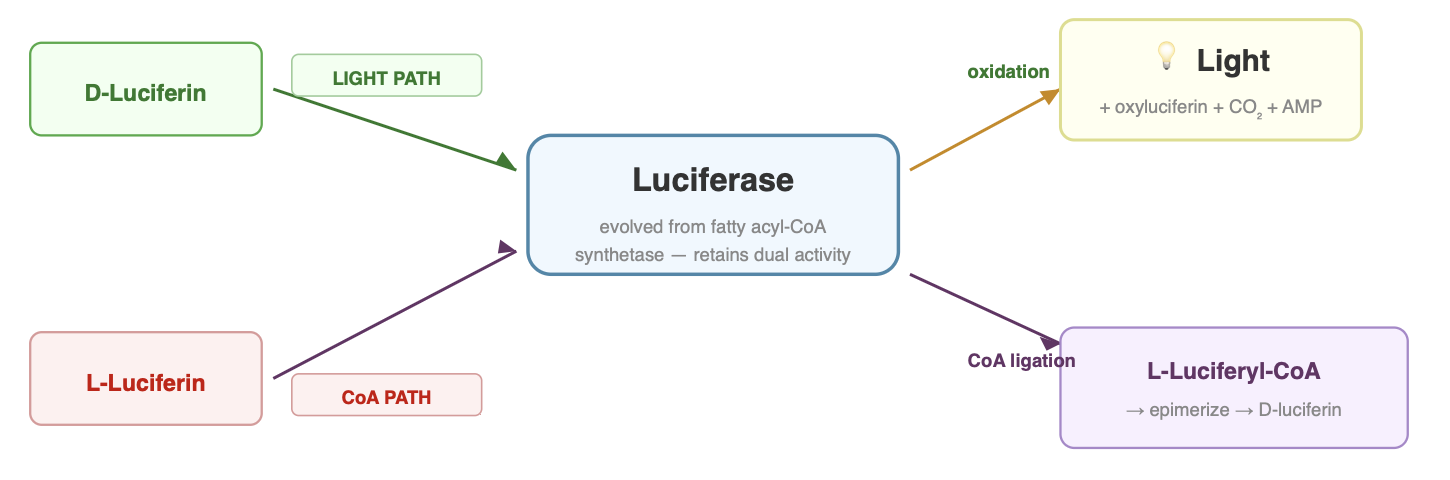
- Enantioselective thioesterification: Luciferase selectively converts L-luciferin to L-luciferyl-CoA using ATP and Coenzyme A. The enzyme treats the two enantiomers differently: D-luciferin is directed into the light-producing reaction, while L-luciferin can be diverted into the CoA-ligase pathway.
- Epimerization: L-luciferyl-CoA spontaneously epimerizes (flips its stereochemistry) to D-luciferyl-CoA through enol formation. This step is non-enzymatic. The CoA thioester makes the adjacent chiral center acidic enough to racemize on its own.
- Thioester hydrolysis: A thioesterase (ACOT-type enzyme) cleaves the CoA off, releasing free D-luciferin ready for the bioluminescence reaction. The specific thioesterase in fireflies has not been definitively identified, though ACOT1 is a strong candidate.
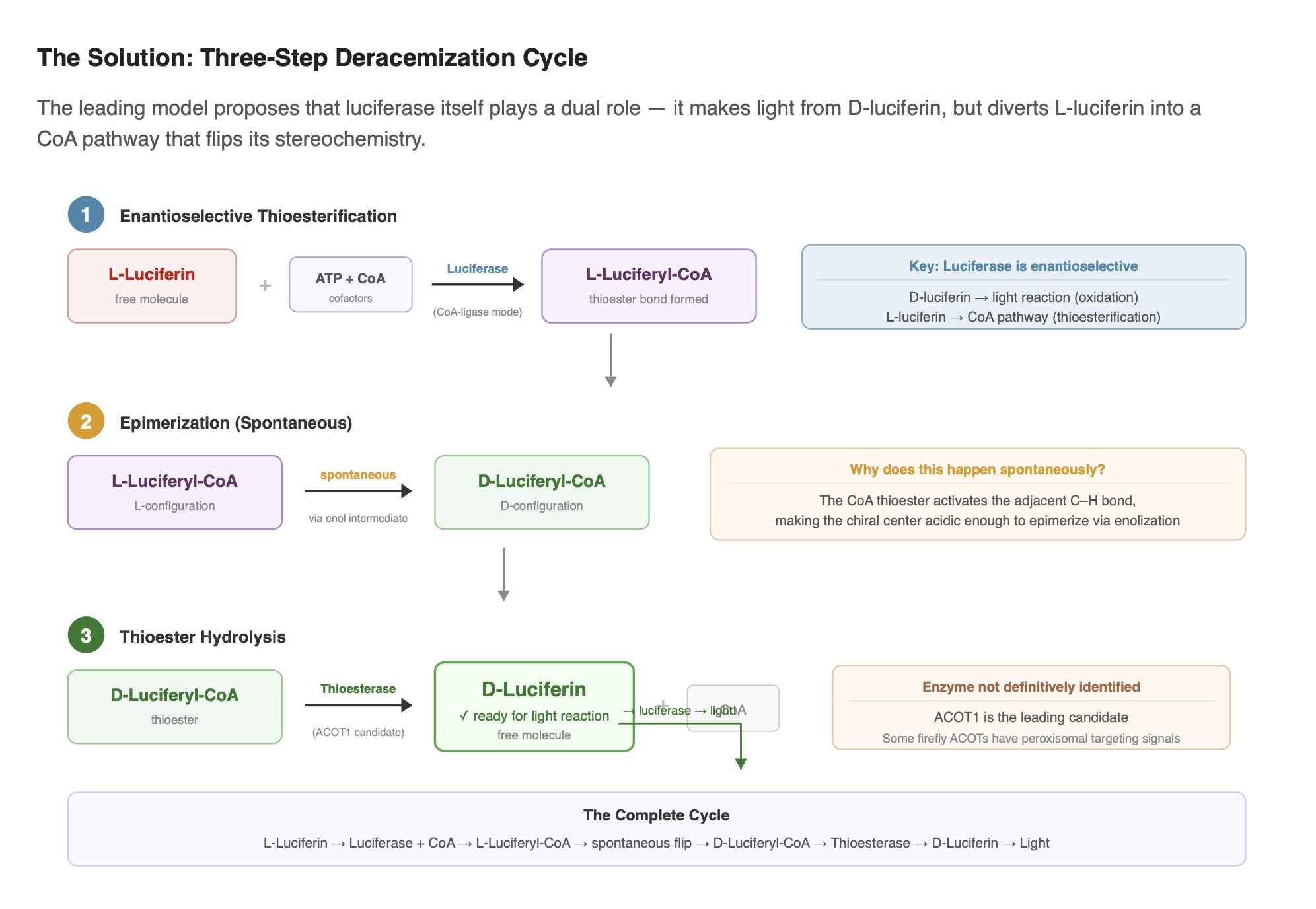
This concept connects back to the enzyme behavior described earlier in the Core Reaction section, where luciferase shows different catalytic handling of the two enantiomers.
Dictionary (9 terms)
- Enantiomer
- One of two mirror-image forms of a molecule that cannot be superimposed on each other, like left and right hands.
- D- and L-
- Prefixes denoting the two mirror-image configurations at a chiral center; firefly luciferase only produces light with D-luciferin.
- Deracemization
- A process that converts a mixture of both enantiomers (a racemic mixture) into predominantly one enantiomer.
- Enantioselective
- A reaction or enzyme that preferentially acts on one enantiomer over the other.
- Thioesterification
- Forming a thioester bond (C=O bonded to sulfur) between a molecule and Coenzyme A.
- Epimerization
- Flipping the stereochemistry at a single chiral center in a molecule, converting one enantiomer to the other.
- Coenzyme A (CoA)
- A common cellular cofactor that forms thioester bonds with various molecules; acts as an "activating handle" for metabolic reactions.
- Thioesterase
- An enzyme that cleaves thioester bonds, releasing the molecule from its CoA handle.
- ACOT1
- Acyl-CoA thioesterase 1; a candidate enzyme for the final hydrolysis step in the luciferin deracemization pathway.
Sulfation and storage
Fireflies do not necessarily use all of their luciferin immediately. A luciferin sulfotransferase (LST) can add a sulfate group to the 6-hydroxyl, producing sulfoluciferin. This sulfated form is inactive, meaning luciferase cannot use it as a substrate. Because of this, it is thought to function as a storage or regulatory form that helps control how much active luciferin is available at any given time.
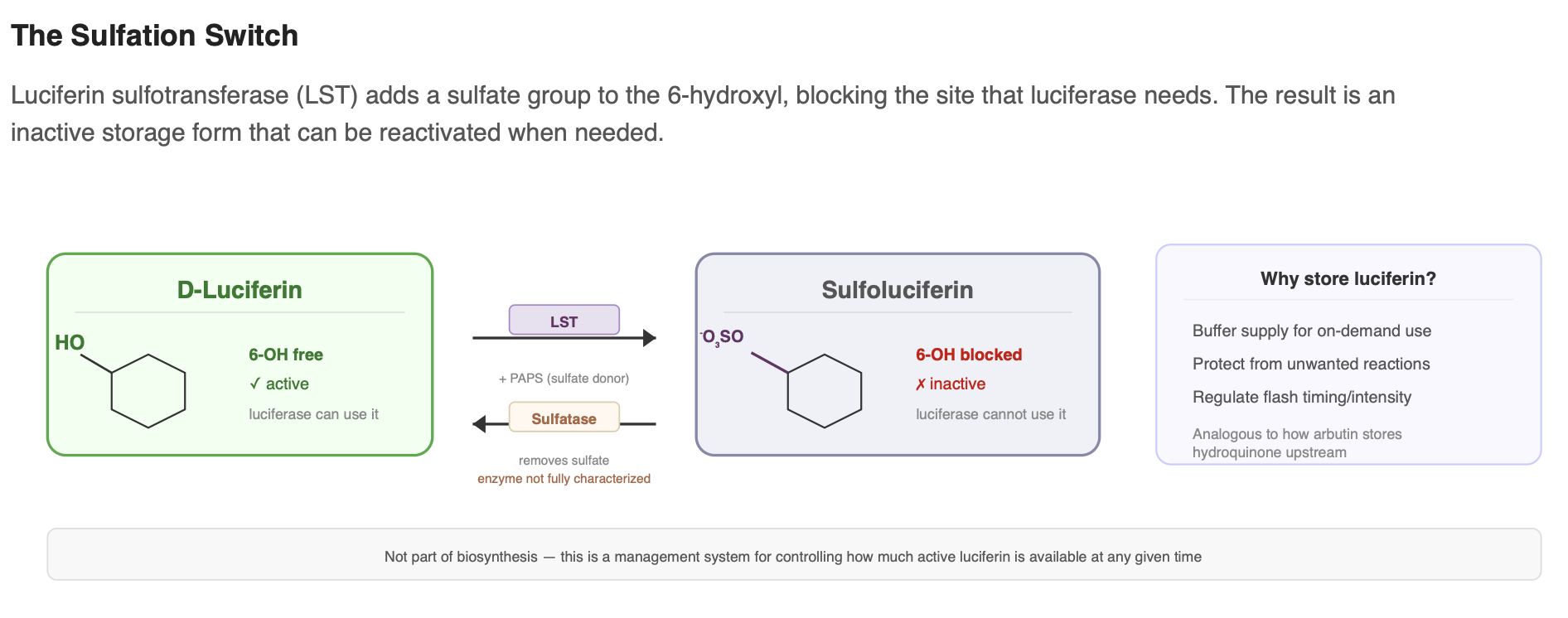
When luciferin is needed, the sulfate group can likely be removed by a sulfatase, releasing free, active luciferin again. While the exact enzyme responsible for this step has not been fully characterized, this reversible sulfation system appears to be part of how the firefly manages and buffers its luciferin supply. This process is not strictly part of biosynthesis, but it is an important part of how the organism regulates luciferin availability.
Recycling: oxyluciferin to luciferin?
After light emission, the spent product is oxyluciferin. It would be advantageous if fireflies could recycle this product rather than relying entirely on de novo luciferin synthesis. A “luciferin-regenerating enzyme” (LRE) was reported to convert oxyluciferin into 2-cyano-6-hydroxybenzothiazole (CHBT), a compound that can condense with cysteine to regenerate luciferin.
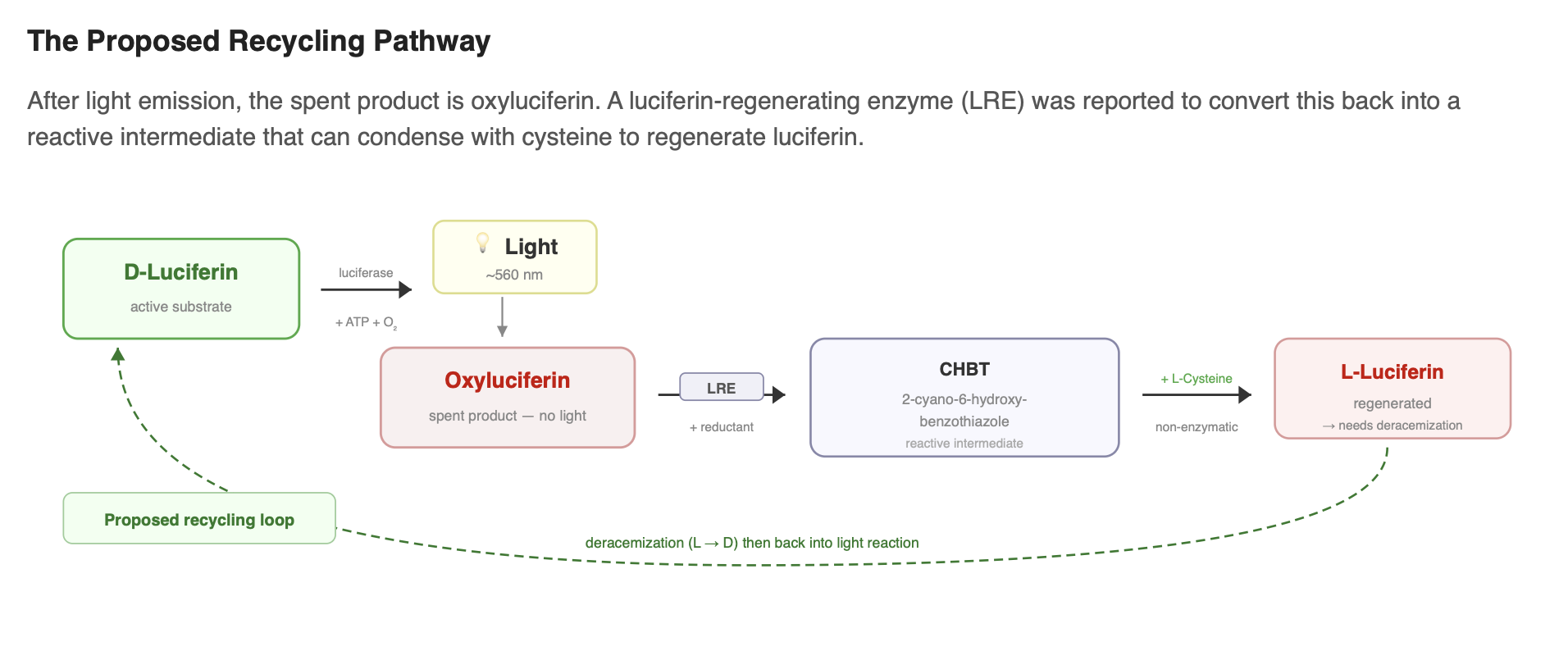
However, the physiological importance of this recycling route remains debated. LRE has been suggested to lack a canonical peroxisomal targeting signal, raising questions about whether it localizes to the same compartment as luciferase, and multiple reviews note that LRE’s in-vivo role still requires clarification.
In short, LRE supports a plausible recycling mechanism in vitro, but whether fireflies recycle oxyluciferin efficiently in vivo, and by what exact enzyme set, remains an open question.
Dictionary (5 terms)
- Phenolate
- The deprotonated form of a phenol (–OH → –O⁻); carries a negative charge and is a stronger electron donor into an aromatic ring.
- Luciferin sulfotransferase (LST)
- An enzyme that adds a sulfate group to luciferin’s 6-hydroxyl, producing inactive sulfoluciferin for storage.
- Luciferin regenerating enzyme (LRE)
- A proposed enzyme that converts spent oxyluciferin back into a benzothiazole intermediate for recycling; its role in vivo is debated.
- 2-Cyano-6-hydroxybenzothiazole (CHBT)
- A proposed intermediate in the recycling pathway; condenses spontaneously with cysteine to form luciferin.
- Peroxisomal targeting sequence
- A short amino acid signal (typically -SKL at the C-terminus) that directs a protein to be imported into peroxisomes.
Compartmentalization: where it all happens
The chemistry described above does not occur randomly throughout the cell. In fireflies, bioluminescence is spatially organized, and understanding that organization matters for anyone trying to recreate the system in another organism.
The lantern organ
Firefly bioluminescence occurs in a specialized organ called the lantern, located on the ventral abdomen. The lantern contains light-producing cells called photocytes, arranged in rosettes around oxygen-delivering tubes called tracheoles. Luciferin biosynthesis, the light reaction, and luciferin management processes are thought to be largely concentrated in these cells.
The peroxisome
Within each photocyte, the light-producing reaction takes place inside peroxisomes, small membrane-bound organelles (Keller et al. 1987). Firefly luciferase has a C-terminal peroxisomal targeting signal (the tripeptide –SKL), which directs it into peroxisomes after being synthesized in the cytosol (Gould et al. 1987). Photocytes are densely packed with peroxisomes, and these organelles are the primary sites where luciferase operates.
The peroxisome likely does more than just contain the reaction. Concentrating enzymes and substrates in a small space can increase effective local concentrations and improve reaction efficiency. The peroxisomal environment may also help provide favorable redox conditions and pH for luciferase activity.
Some candidate enzymes involved in luciferin management may also localize to peroxisomes. For example, certain ACOT-type thioesterases carry predicted PTS1 signals (Zhang et al. 2020), suggesting that parts of the L→D conversion cycle could occur inside this compartment.
Multiple compartments
Not all pathway components appear to be peroxisomal. The proposed recycling enzyme LRE lacks a clear peroxisomal targeting signal, suggesting it may function in the cytosol. Enzymes involved in earlier steps, such as β-glucosidase (BGL) and laccase, are also likely to operate outside the peroxisome. This implies that luciferin biosynthesis may span multiple cellular compartments, with earlier chemical steps occurring in the cytosol and the final light-producing reaction occurring in peroxisomes.
How fireflies control their flash
The on/off switching of firefly light is controlled largely by oxygen availability. Neural signals trigger nitric oxide (NO) release in the lantern (Trimmer et al. 2001). NO temporarily inhibits mitochondrial respiration, reducing oxygen consumption by mitochondria and allowing more oxygen to reach the peroxisomes. This influx of oxygen enables the luciferase reaction and produces a flash. When NO signaling stops, mitochondria resume consuming oxygen, reducing peroxisomal oxygen levels and turning the light off (Aprille et al. 2004). In effect, the flash is controlled by regulating oxygen access.
Why this matters for plant engineering
Plants already contain peroxisomes, where processes like fatty acid β-oxidation and photorespiration occur. The PTS1 import system is highly conserved across eukaryotes, which is why firefly luciferase expressed in plant cells correctly localizes to plant peroxisomes without modification.
For an engineered system, directing luciferase (and possibly certain downstream enzymes) to peroxisomes using their native targeting signals could help recreate the concentrated reaction environment found in firefly photocytes. Enzymes involved in upstream steps may not require targeting, since their small-molecule substrates can likely diffuse between compartments.
The key engineering question is whether compartmentalization significantly affects brightness. In the firefly, the peroxisomal concentration effect clearly contributes to efficiency. In a plant, where enzyme levels may be lower and cells are not specialized for light production, spatial organization could be an important factor in achieving visible glow.
The other glowing plant: why not use the mushroom system?
There is already a glowing plant on the market. In 2020, Mitiouchkina et al. demonstrated autonomously bioluminescent tobacco using the fungal bioluminescence pathway from Neonothopanus nambi (Mitiouchkina et al. 2020), and the company Light Bio now sells a glowing petunia commercially. That system works by tapping into caffeic acid, a metabolite plants already produce abundantly, and converting it through a four-enzyme pathway into a fungal luciferin that emits green light.
So why not just use that?
The firefly system is fundamentally brighter. Firefly bioluminescence has a quantum yield of roughly 41% (Ando et al. 2008), meaning 41 out of every 100 reactions produce a photon. That is the highest of any known bioluminescence system. The fungal system is significantly lower. On a per-reaction basis, firefly chemistry converts more energy into light and less into heat.
The emission wavelength also matters. Firefly luciferase emits at ~560 nm, which sits near the peak of human dark-adapted vision. The fungal system emits a dimmer, greener light. In practice, Light Bio’s Firefly Petunia produces what’s been described as a gentle glow comparable to moonlight; visible in a dark room, but not dramatically so, and most promotional photos use long camera exposures.
The firefly pathway is harder to engineer. The substrate problem that this entire blog post describes is real and unsolved. But if it works, the theoretical brightness ceiling is higher. That’s the bet: a harder engineering challenge in exchange for a brighter result.
Part II
Plant Engineering
Everything above describes the biology as it exists in fireflies.
What follows is about recreating it in plants.
The engineering plan: four genes to close a 40-year gap
Since Keith Wood showed in 1986 that plants expressing firefly luciferase glow when watered with luciferin (Ow et al. 1986), the goal has been obvious: make the plant produce its own luciferin so it can glow autonomously. That requires giving the plant the enzymatic machinery for the biosynthetic pathway described above.
Plants already have most of what they need. They make quinones and hydroquinones through normal metabolism. They have abundant L-cysteine. They have ATP, O₂, Mg²⁺, and functioning peroxisomes with a conserved PTS1 import system. What they’re missing are the specific enzymes that channel those precursors into luciferin and then into light.
My project aims to supply those missing pieces with four genes:
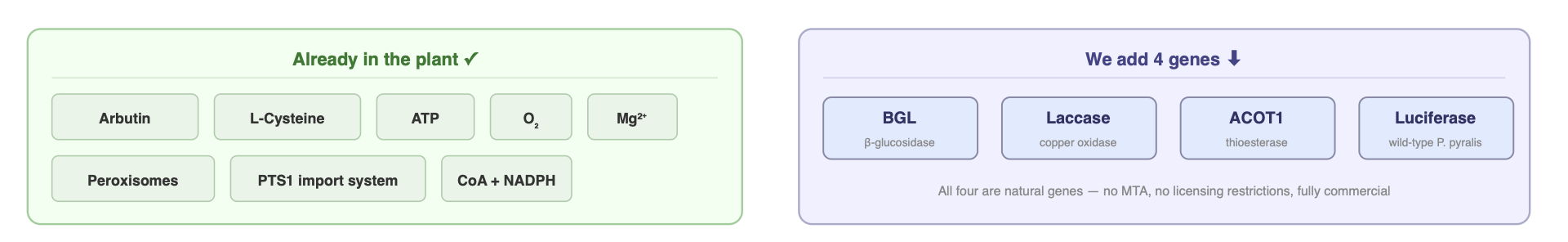
1. BGL (β-glucosidase)
Role: Release the precursor from storage
As described in the biosynthesis section, the hydroquinone/quinone precursor is proposed to be stored as arbutin (Oba et al. 2013), hydroquinone locked behind a glucose molecule in a safe, less reactive glycoside form. BGL cleaves that glucose off, releasing free hydroquinone into the pathway. Certain species have natural arbutin stores, and plant-engineering studies have also built hydroquinone supply routes in hosts that do not. This step can act as a key gate for pathway flux: without effective β-glucosidase activity, much of the precursor remains sequestered and downstream conversion is strongly limited. Plants do have endogenous β-glucosidases, but a firefly-derived BGL helps ensure the needed substrate specificity and expression level.
A plant is very likely to successfully express a β-glucosidase (BGL) gene, because plants already produce many similar enzymes and generally tolerate them well. The main uncertainty is not expression but usefulness. BGL will only help if the plant actually contains arbutin or related glycosides to release hydroquinone from. If that precursor pool is present and accessible, BGL can act as a gate that increases pathway flux. If little or no arbutin exists in the host species, the enzyme may have minimal effect. So the biological acceptance risk is low, but the impact depends heavily on the plant’s existing chemistry.
2. Laccase
Role: Oxidize hydroquinone to the reactive quinone
Free hydroquinone likely needs to be oxidized to p-benzoquinone before it can condense with cysteine. Laccases (copper-containing oxidase enzymes) are strong candidates to carry out this conversion. This same enzyme class has been shown to enhance luciferin-forming chemistry in vitro (de Souza et al. 2022), suggesting it could play a dual role: generating the quinone and promoting the initial condensation step. Plants already contain many endogenous laccases, but including a pathway-specific laccase helps ensure sufficient and reliable oxidation of the precursor.
A plant is very likely to tolerate and express a laccase, since plants already have large native laccase families and are used to this type of enzyme. The main uncertainty is functional, not biological. Laccases oxidize many phenolic molecules, not just hydroquinone, so they could help generate the needed quinone precursor but might also oxidize other plant compounds or act in the wrong compartment. Overall, expression risk is low; usefulness depends on localization and how much precursor is present.
3. ACOT1 (acyl-CoA thioesterase)
Role: Help complete the deracemization cycle
This enzyme is intended to close the stereochemical inversion loop (Niwa et al. 2006; Zhang et al. 2020). As described in the deracemization section, luciferase can divert L-luciferin into L-luciferyl-CoA, which can spontaneously epimerize to D-luciferyl-CoA. However, this CoA-bound form cannot participate in the light reaction until the CoA group is removed.
ACOT-type thioesterases are strong candidates for this step. They cleave CoA thioesters to release the free acid, which in this case would regenerate free D-luciferin. If this hydrolysis step is inefficient, the cycle could become trapped in CoA-bound intermediates, limiting the amount of usable D-luciferin and reducing light output over time.
ACOT1 is therefore included as a practical way to ensure that any D-luciferyl-CoA formed is converted back into free D-luciferin. It may be the least glamorous gene in the cassette, but it could be important for sustaining continuous light production.
Biologically, the risks to the integrity of the plant are low. Plants already have many native acyl-CoA thioesterases and regularly handle CoA-bound intermediates, so expressing an additional ACOT-type enzyme is unlikely to be toxic or disruptive on its own. It doesn’t consume much energy, doesn’t create reactive byproducts, and acts on small molecules the cell is used to processing.
The main uncertainty is not plant health but usefulness. It’s still unclear how much the luciferin pathway actually depends on the CoA deracemization loop, and plants may already have endogenous thioesterases that can perform the same function. So ACOT1 is unlikely to harm the plant, but its impact on light output could range from essential to redundant depending on how the pathway behaves in practice.
4. Wild-type Photinus pyralis luciferase
Role: The light reaction itself
Wild-type Photinus pyralis luciferase is the best fit for a plant engineered to produce its own D-luciferin because it is naturally optimized for that exact substrate. Unlike engineered variants such as AkaLuc, which were designed for synthetic luciferins and perform worse with native D-luciferin, the original enzyme efficiently converts D-luciferin into light. It has a very high quantum yield (Ando et al. 2008), emits yellow-green light (~560 nm) near the peak sensitivity of human night vision, and already contains a built-in peroxisomal targeting signal (–SKL) that correctly localizes it inside eukaryotic cells (Gould et al. 1987). It is also widely used, well-characterized, and free of licensing restrictions. For a system built around endogenous D-luciferin production, it’s not a fallback option, it’s the most compatible and practical choice.
The honest picture: a minimum viable pathway
These four genes represent a minimum viable pathway. The smallest gene set that could, in principle, produce autonomous bioluminescence. The logic is:
BGL and laccase supply the reactive quinone from stored precursors. The condensation with cysteine may proceed spontaneously or with laccase assistance (it works non-enzymatically in vitro, albeit at low yield; Kanie et al. 2016). The benzothiazoline-to-benzothiazole oxidation may happen through air oxidation, since the molecule thermodynamically wants to become aromatic. ACOT1 closes the deracemization loop, and wild-type luciferase handles the light reaction.
But this is a hypothesis, not a guarantee. The pathway may be missing pieces. Candidates for additional genes include a dedicated condensase for the quinone-cysteine reaction (no one has identified one, but the low non-enzymatic yield suggests one may exist), a hydroxylase such as PPYR_02911 if the 6-OH isn’t inherited from the quinone precursor, and possibly a dedicated oxidase for the aromatization step.
If the four-gene plant glows, that’s a significant result. It would suggest that the minimal pathway is sufficient. If it doesn’t, the debugging process is still scientifically valuable. Each failure point helps narrow the search for missing enzymes and clarifies how the natural system actually works.
Either outcome advances understanding. That’s what makes this a real experiment rather than just an engineering exercise.
Living section · Last updated Feb 2026
Candidates from the lantern: genes that could complete the pathway
My four-gene system is designed as a minimum viable pathway, but the firefly almost certainly uses additional enzymes that haven’t all been identified. This section will evolve alongside the experiments, with new candidate genes added as stronger evidence emerges.
PPYR_02911 Cytochrome P450, CYP4C family
My top current candidate. It shows a lantern TPM of 534, is 21× upregulated, and has the most statistically significant differential expression of the group (q = 8.45e-20). It sits in a tandem duplication with PPYR_02910. The two genes are 87% identical and only 4 kb apart, a classic signature of gene neofunctionalization, where one copy evolves a new function while the other maintains the original role. BLAST results are also striking: the closest matches are almost exclusively from bioluminescent species, with the first non-luminous beetle appearing only around 49% identity. This expression and evolutionary profile is consistent with an enzyme that may have specialized for the bioluminescence system. A plausible role is hydroxylation of the benzothiazole ring at the 6-position, if that oxygen is not inherited directly from the quinone precursor. To my knowledge, this gene has not been widely discussed as a luciferin-pathway candidate.
PPYR_14056 4-Coumarate:CoA ligase
Previously proposed as a candidate (Zhang et al. 2020). It shows strong lantern enrichment and has chemistry consistent with CoA-linked activation steps. This could connect to the proposed deracemization loop or to broader CoA-mediated processing of aromatic precursors, potentially interacting with peroxisomal metabolism.
PPYR_14756 UDP-glycosyltransferase
The strongest statistical signal of the group (q = 2.45e-24). This enzyme is more likely to serve a support or regulatory role than a core biosynthetic one. Glycosyltransferases often convert reactive small molecules into stable storage forms, similar to how hydroquinone can be stored as arbutin. This gene could be involved in temporarily glycosylating luciferin or its precursors to control availability and reduce toxicity.
Citations
- Conti, E., Franks, N.P., & Brick, P. (1996). “Crystal structure of firefly luciferase throws light on a superfamily of adenylate-forming enzymes.” Structure, 4(3):287–298.
- Inouye, S. (2010). “Firefly luciferase: an adenylate-forming enzyme for multicatalytic functions.” Cellular and Molecular Life Sciences, 67:387–404.
- de Wet, J.R., Wood, K.V., Helinski, D.R., & DeLuca, M. (1985). “Cloning of firefly luciferase cDNA and the expression of active luciferase in Escherichia coli.” Proceedings of the National Academy of Sciences, 82(23):7870–7873.
- Ow, D.W., Wood, K.V., DeLuca, M., de Wet, J.R., Helinski, D.R., & Howell, S.H. (1986). “Transient and stable expression of the firefly luciferase gene in plant cells and transgenic plants.” Science, 234(4778):856–859.
- Oba, Y., Yoshida, N., Kanie, S., Ojika, M., & Inouye, S. (2013). “Biosynthesis of firefly luciferin in adult lantern: decarboxylation of ʟ-cysteine is a key step for benzothiazole ring formation in firefly luciferin synthesis.” PLoS ONE, 8(12):e84023.
- Kanie, S., Nishikawa, T., Ojika, M., & Oba, Y. (2016). “One-pot non-enzymatic formation of firefly luciferin in a neutral buffer from p-benzoquinone and cysteine.” Scientific Reports, 6:24794.
- Kanie, S., Nakai, R., Ojika, M., & Oba, Y. (2018). “2-S-cysteinylhydroquinone is an intermediate for the firefly luciferin biosynthesis that occurs in the pupal stage of the Japanese firefly, Luciola lateralis.” Bioorganic Chemistry, 80:223–229.
- de Souza, D.R., Silva, J.R., Moreira, A., & Viviani, V.R. (2022). “Biosensing firefly luciferin synthesis in bacteria reveals a cysteine-dependent quinone detoxification route in Coleoptera.” Scientific Reports, 12:14815.
- Zhang, R., et al. (2020). “Genomic and experimental data provide new insights into luciferin biosynthesis and bioluminescence evolution in fireflies.” Scientific Reports, 10:15882.
- Niwa, K., Nakamura, M., & Ohmiya, Y. (2006). “Stereoisomeric bio-inversion key to biosynthesis of firefly D-luciferin.” FEBS Letters, 580(22):5283–5287.
- Keller, G.A., Gould, S., DeLuca, M., & Subramani, S. (1987). “Firefly luciferase is targeted to peroxisomes in mammalian cells.” Proceedings of the National Academy of Sciences, 84(10):3264–3268.
- Gould, S.J., Keller, G.A., & Subramani, S. (1987). “Identification of a peroxisomal targeting signal at the carboxy terminus of firefly luciferase.” Journal of Cell Biology, 105(6):2923–2931.
- Trimmer, B.A., Aprille, J.R., Dudzinski, D.M., Lagace, C.J., Lewis, S.M., Michel, T., Qazi, S., & Zayas, R.M. (2001). “Nitric oxide and the control of firefly flashing.” Science, 292(5526):2486–2488.
- Aprille, J.R., Lagace, C.J., Modica-Napolitano, J., & Trimmer, B.A. (2004). “Role of nitric oxide and mitochondria in control of firefly flash.” Integrative and Comparative Biology, 44(3):213–219.
- Mitiouchkina, T., et al. (2020). “Plants with genetically encoded autoluminescence.” Nature Biotechnology, 38:944–946.
- Ando, Y., Niwa, K., Yamada, N., Enomoto, T., Irie, T., Kubota, H., Ohmiya, Y., & Akiyama, H. (2008). “Firefly bioluminescence quantum yield and colour change by pH-sensitive green emission.” Nature Photonics, 2:44–47.
- Seliger, H.H. & McElroy, W.D. (1959). “Quantum yield in the oxidation of firefly luciferin.” Biochemical and Biophysical Research Communications, 1:21–24.
- Seliger, H.H. & McElroy, W.D. (1960). “Spectral emission and quantum yield of firefly bioluminescence.” Archives of Biochemistry and Biophysics, 88:136–141.